-
Tematy
-
Odpowiedzi
-

AnT 79
-
AnT 79
Minęły dwa miesiące od wsadzenia nowej konstrukcji i wsypaniu podłoża . Rzecz jasna wiadome - zawsze trzeba coś spier.... i . Wymieniłem konstrukcje wyjąłem korale ,ale za to nie zmniejszyłem światła , nie wziąłem poprawki na ballingu , do tego dalej podlewałem namiętnie aminio . Kapnąłem się jak zaczęło robić się za zielono , nie taki był plan . Wiec szybka korekta . Ale do wody poszło co miało pójść wiec - co zostało - CZAS . I powoli się prostuje .Nalot który się pokazał zanika powoli . Cjano ,dino ,okrzemków - brak / niebyło . Wczoraj nawet mandaryny miały zaloty . siały ikrą w toń .









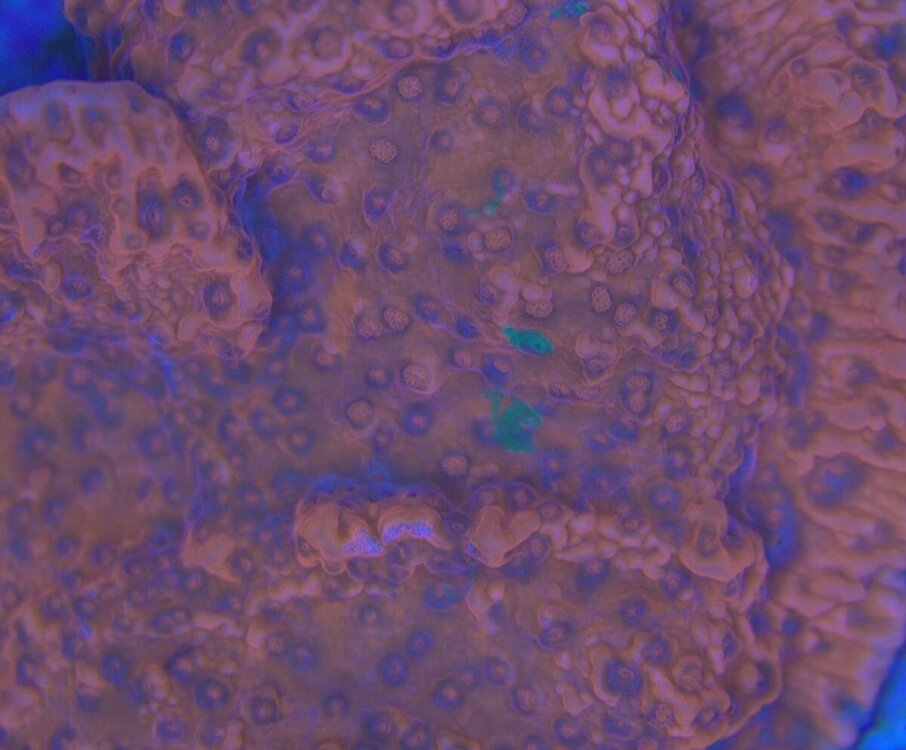
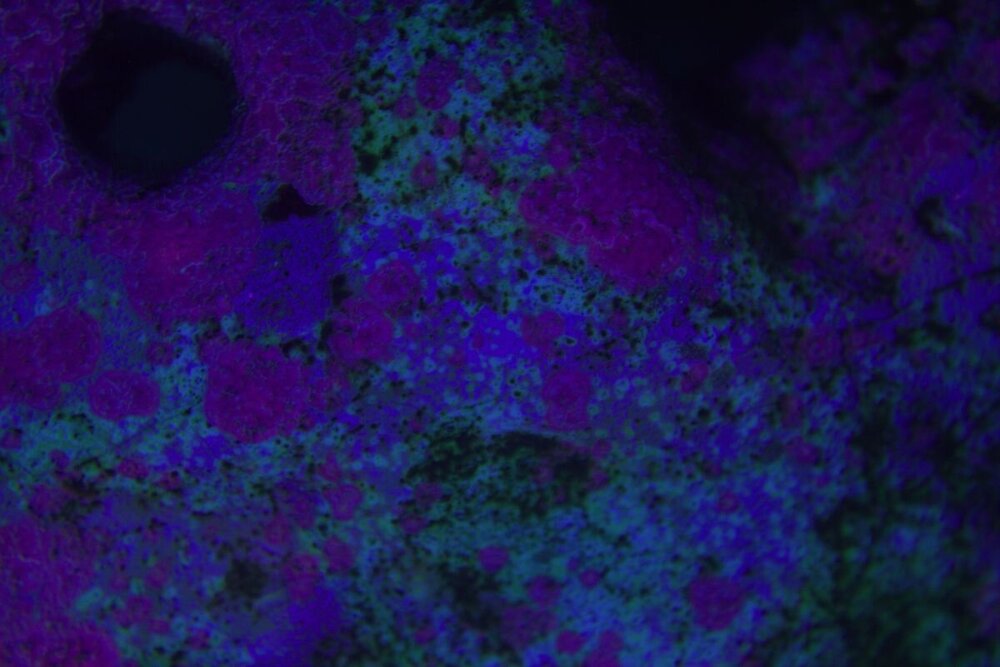
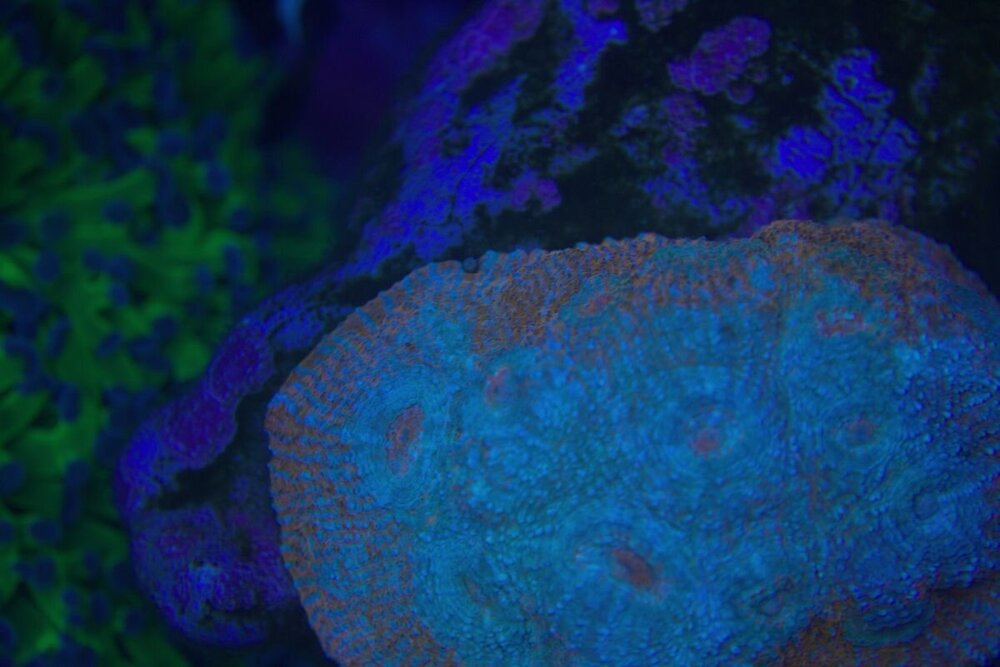
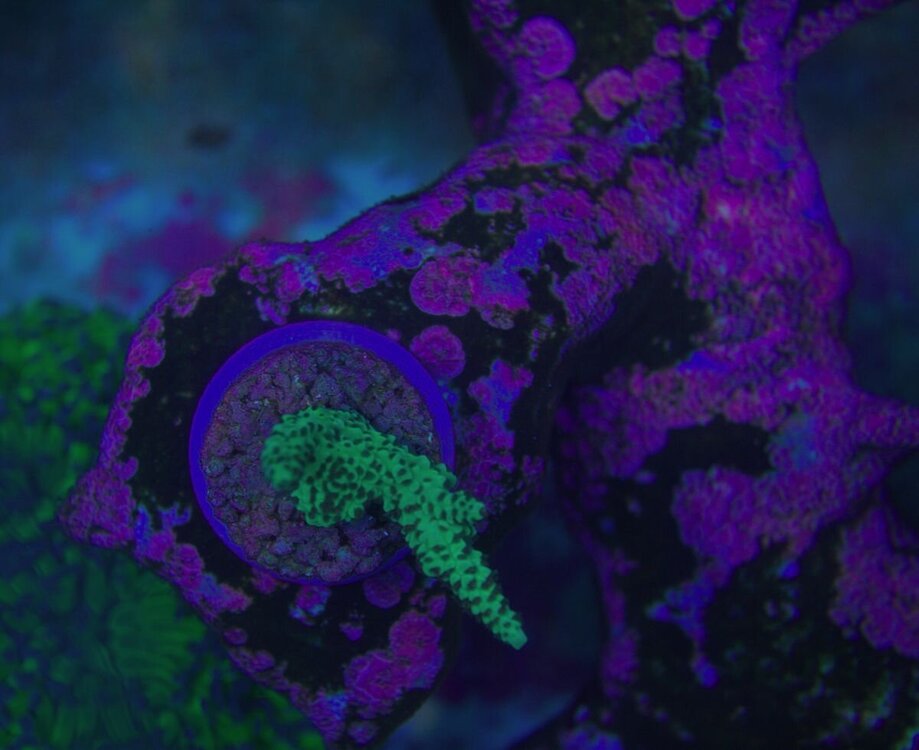
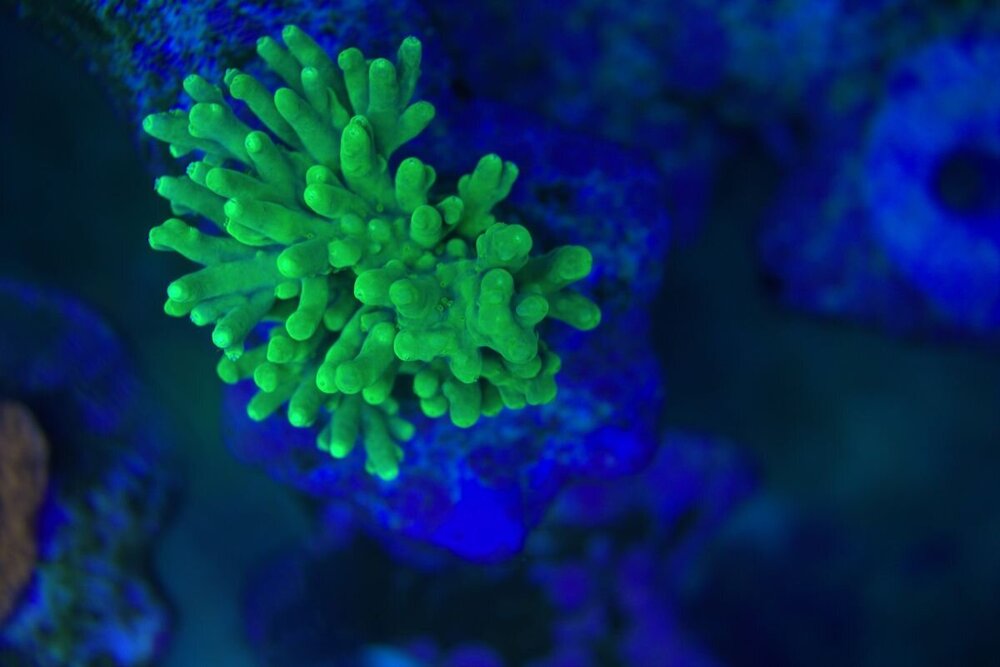

-
AnT 79
W dniu 7.01.2026 o 14:15, Atixure napisał:Cześć,
Na start planuję tanią kostkę od Zenona (nano), bez szaleństw — podstawy, nauka, spokojne tempo.
Witam ,ale już wybrałeś co chcesz mieć to trzeba u niego podglądać lub
Tania Kostka Akwarium Morskie Zenona SET 3 + Skała + gratisy Sklep akwarystyczny online ▷ Trzmiel
Wiec ...
-

Atixure 0
Cześć,
chcę zacząć przygodę z akwarystyką morską. Na start planuję tanią kostkę od Zenona (nano), bez szaleństw — podstawy, nauka, spokojne tempo.
Szukam rad od bardziej doświadczonych:
• co faktycznie trzeba dokupić na start,
• na co uważać przy pierwszym zalewaniu,
• czego NIE robić, żeby nie utopić kasy i zwierzaków.
Będę wdzięczny za konkretne wskazówki i sprawdzone patenty.
Pozdro

Wysłane z iPhone za pomocą Tapatalk
-
AnT 79
Dziś.

Taka ciekawostka -mat po prawej , lewa połysk i na nim letki nalot zielony, ślimaki bez problemu dają radę. Więc parę dni i powinno być pozytywnie . Osad krzemianowy z piasku praktyczne zniknął.
-

















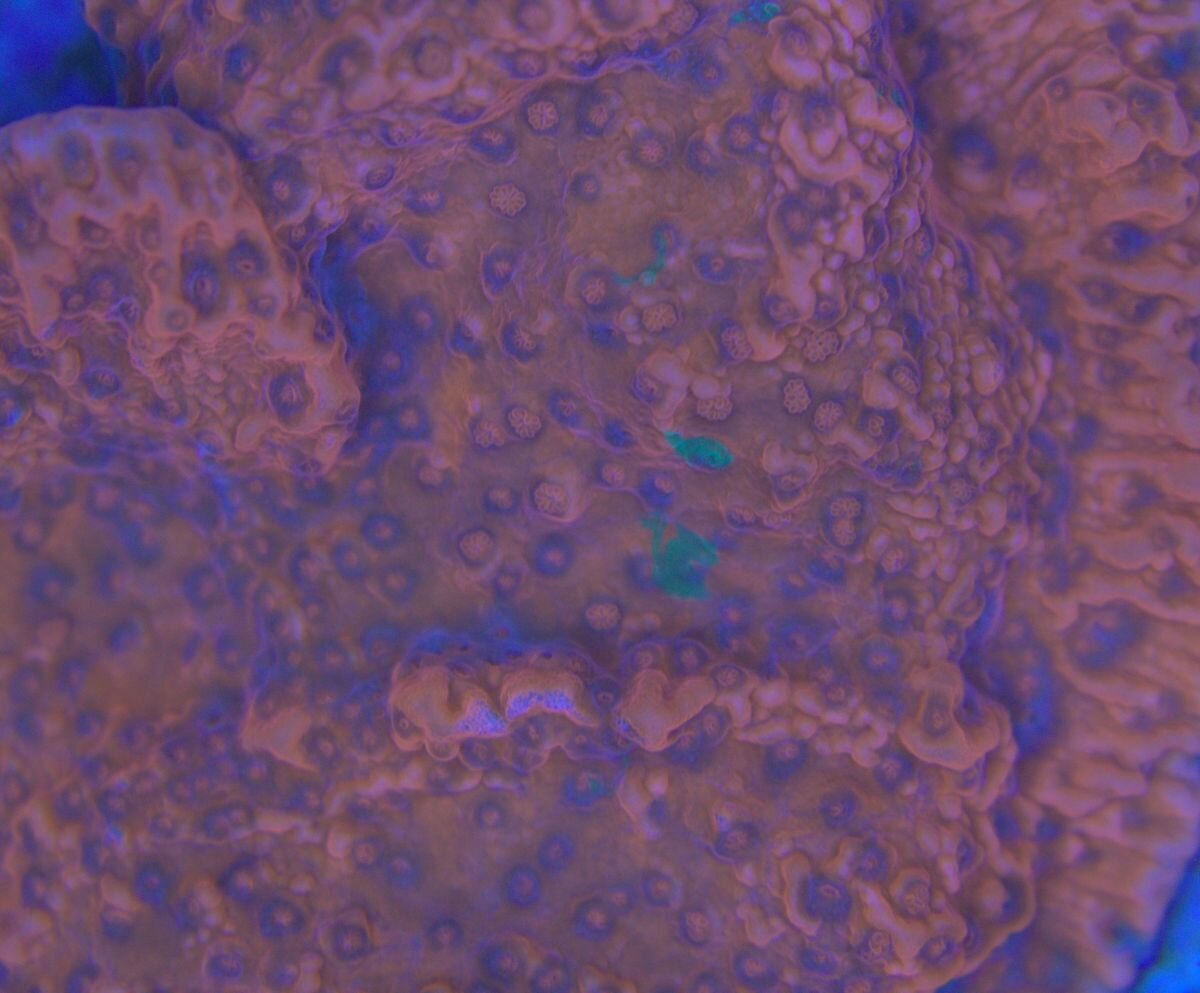
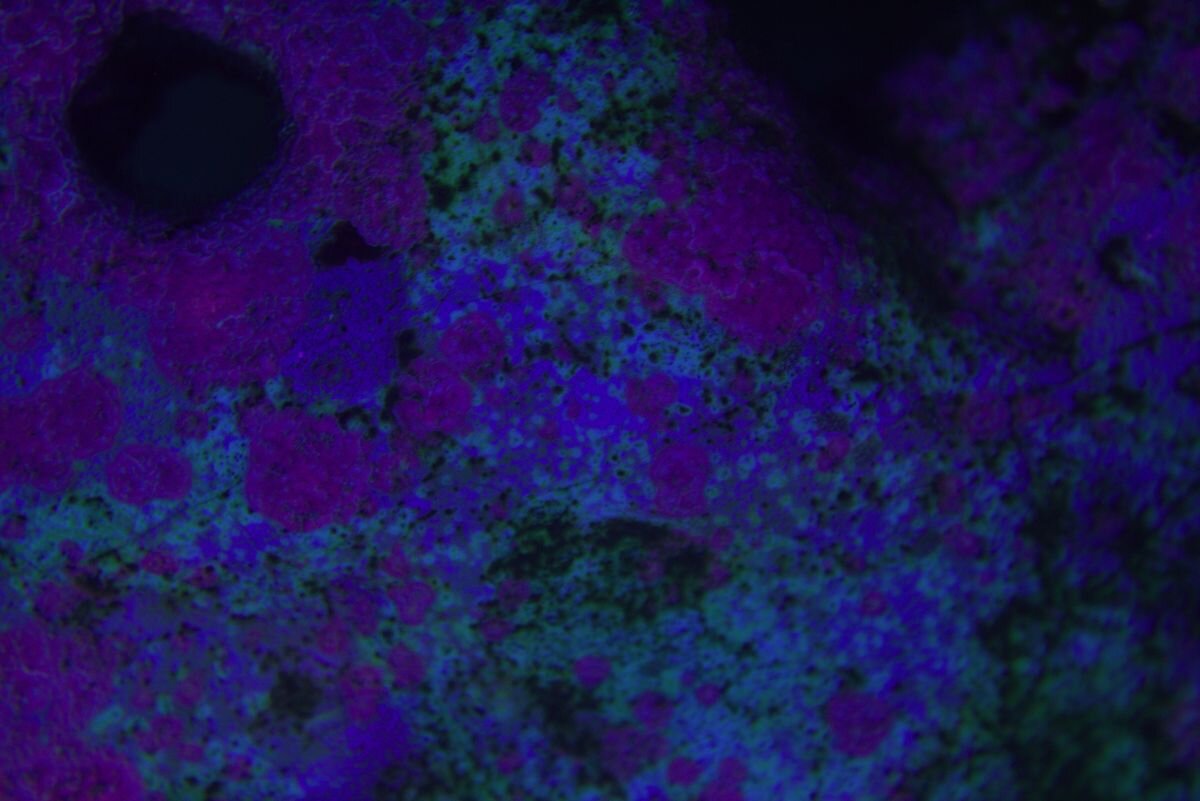
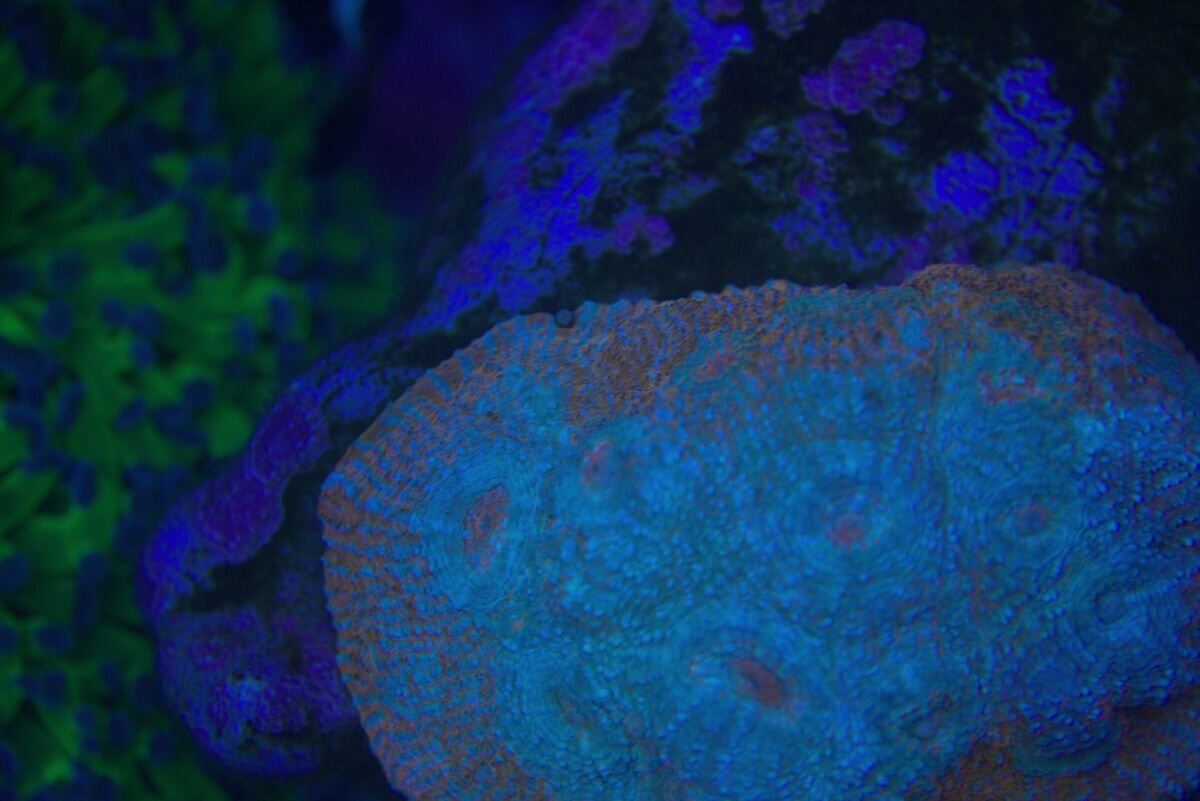


Cała aktywność
Kanał aktualizowany automatycznie
- Ostatni tydzień
-
 Zarejestrował się nowy użytkownik:
Tad
Zarejestrował się nowy użytkownik:
Tad
- Wcześniejsza
-
Powoli Po tych wszystkich akcjach i tak udało się. Ale za to jaka nauka .
-
Minęły dwa miesiące od wsadzenia nowej konstrukcji i wsypaniu podłoża . Rzecz jasna wiadome - zawsze trzeba coś spier.... i . Wymieniłem konstrukcje wyjąłem korale ,ale za to nie zmniejszyłem światła , nie wziąłem poprawki na ballingu , do tego dalej podlewałem namiętnie aminio . Kapnąłem się jak zaczęło robić się za zielono , nie taki był plan . Wiec szybka korekta . Ale do wody poszło co miało pójść wiec - co zostało - CZAS . I powoli się prostuje .Nalot który się pokazał zanika powoli . Cjano ,dino ,okrzemków - brak / niebyło . Wczoraj nawet mandaryny miały zaloty . siały ikrą w toń .
-
Witam ,ale już wybrałeś co chcesz mieć to trzeba u niego podglądać lub Tania Kostka Akwarium Morskie Zenona SET 3 + Skała + gratisy Sklep akwarystyczny online ▷ Trzmiel Wiec ...
-
Cześć, chcę zacząć przygodę z akwarystyką morską. Na start planuję tanią kostkę od Zenona (nano), bez szaleństw — podstawy, nauka, spokojne tempo. Szukam rad od bardziej doświadczonych: • co faktycznie trzeba dokupić na start, • na co uważać przy pierwszym zalewaniu, • czego NIE robić, żeby nie utopić kasy i zwierzaków. Będę wdzięczny za konkretne wskazówki i sprawdzone patenty. Pozdro [emoji112] Wysłane z iPhone za pomocą Tapatalk
-
Dziś. Taka ciekawostka -mat po prawej , lewa połysk i na nim letki nalot zielony, ślimaki bez problemu dają radę. Więc parę dni i powinno być pozytywnie . Osad krzemianowy z piasku praktyczne zniknął.
-
-
Właśnie dodałem jeszcze piasek . I znowu woda mętna, teraz cjano , dino , glony w poczekalni.
-
Konstrukcja zwodowana. Jeszcze piasek , korale takie tam bzdety .
-
Dziś odebrałem , nowa konstrukcja .
-
Nowa zabawka pod aplikacje Tuya - mnie interesuje tylko pomiar ph z dostępem historii . Wiadome sonda do wymiany( czekam ) będzie wiadomo - jedyna godna - Hydrometr . Yieryi WIFI Smart Aquarium PH Meter YY-W9909 Digital PH EC TDS SALT S.G Temp Water Quality Tester Tuya Monitor Control US/EU
-
Podzielę się , oby nie musieliście używać . Jak ktoś będzie miał problem - może sie w ten sposób ratować obsadę . Ja u siebie pierwszy raz odkąd mam akwa użyłem uv - no nie było okazji . Po dwóch tygodniach użycia efekt przyjemny , wcześniej ryby obsypane co niektóre myślałem że już "wieczna rafa" ryby obsypało momentalnie - tak niespodzianka przyniosłem sobie gratis Oprócz intensywnego karmienia różnymi pokarmami doszła uv po paru dniach do sumpa - OCZYWISYTE LAMPA NADAJE SIE TYLKO DO SUMPA ! . Ale za działa na chwile obecna świece 24h na dobę . Tak wiem - lampa tłucze wszystko w przegrodzie co do niej wpada i to dla mnie ważne , nie jak standardowa uv w tubie . Co ciekawe ślimaki nic sobie z tego robią. Wiadomo parametry przy takim ustawieniu czyli - mocne karmienie + tłuczenie uv . Po4 - 0.00 hanka , No3 - 50 - podskoczyło ale to raczej nie problem . Ważne że obsada od trzech tygodni żyje i ma się lepiej . Kto chce może sobie spróbować. Pokolce też przez chwile obsypało ale ładnie zeszło i co fajne cały czas jadły i to ważne . Najgorzej zebrała samica błazenka to już była agonia wręcz ale dała rade mniej jadła przez chwile musiałem wręcz przy karmieniu praktycznie pipetą podawać pod pysk pokarm ale po paru dnia zaczęła sama pobierać . Im więcej różnego pokarmu tym lepiej , dziwne bo robią się bardzo wybredne . Na chwile obecną wraca do normy wszystko .
-
Witam . Jak w temacie używa ktoś dłuższy czas ? Reef Scorpionfish Balling 2l MAX II
-
Wygląda na to, że całkiem długo. Będzie w ogóle?
-
Nowy projekt . Dziś przyszły paczki . Pakowanie pierwsza klasa . Więc: Pierwszy etap jest Ceramika pod specjalne zamówienie @balon Teraz będzie drugi etap , szkliwienie.
-
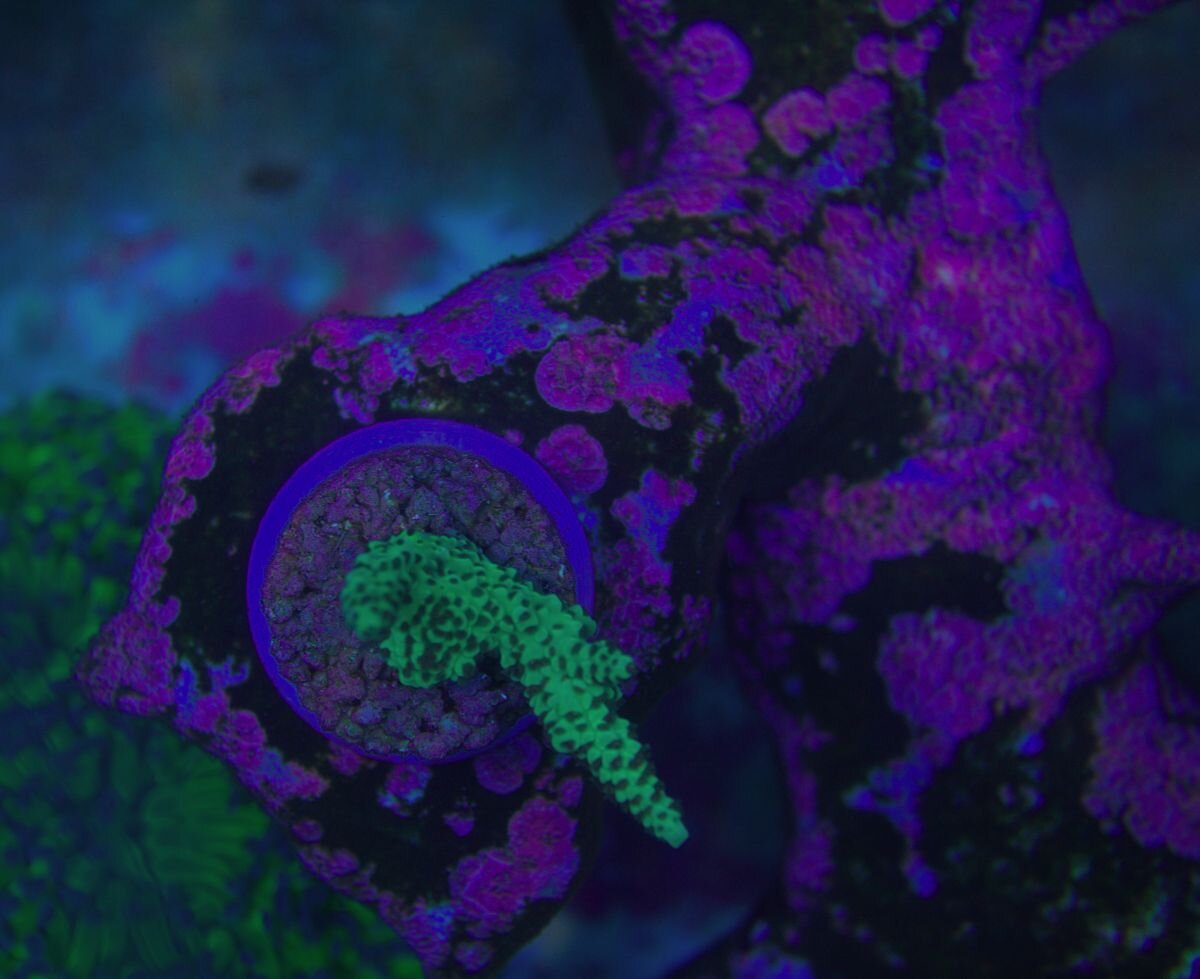
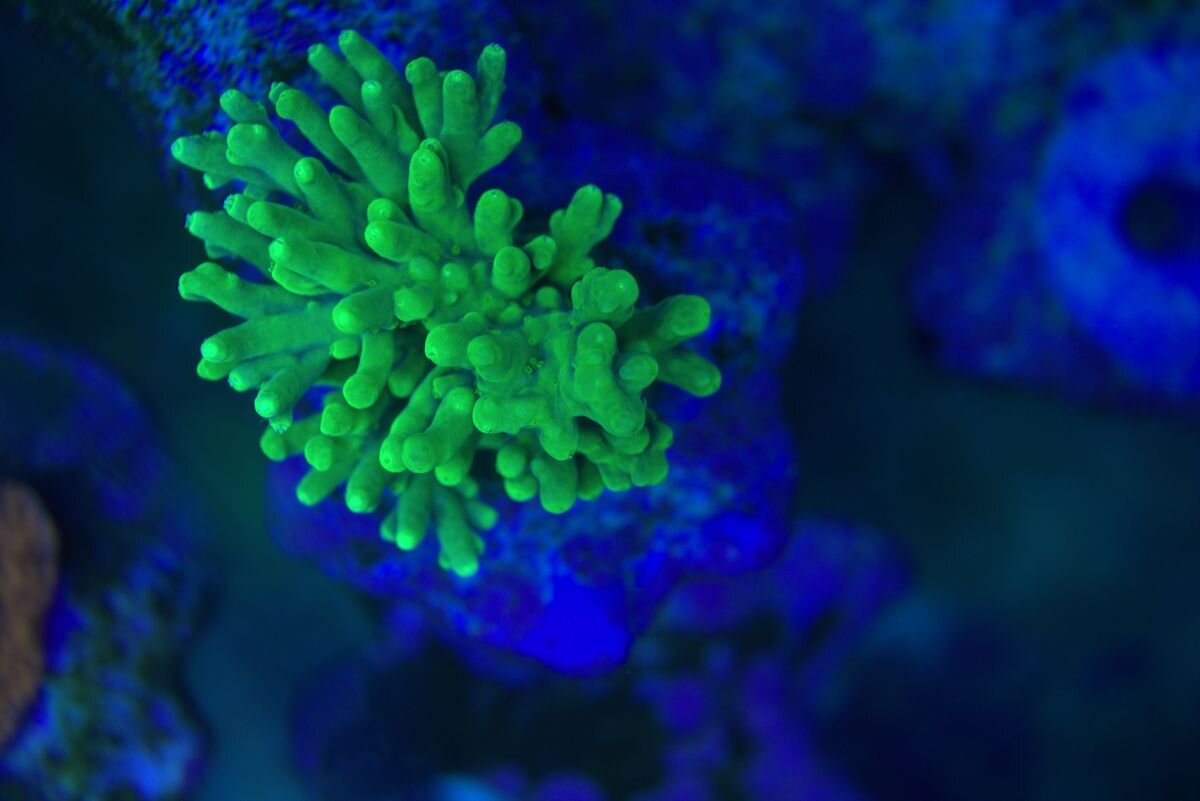
Witam . Daj żyć albo spróbuj dać . Kiedyś kleiłem młodą Fungie do podstawki by nie zginęła w akwa ( za mała ) I niedawno ta sama odpadała młoda Fungia z podstawki i po paru dniach (około półtorej tygodnia ) jest na nowo nowa ,wyrosła z podstawy klejenia . I to jest fajne .Fotki marnej jakości . Ale liczy się efekt . Po lewej pod Naso to co zostało jak odpadła młoda i została przyklejona na nowo , jest z prawej strony .I Foto z dziś jak to wygląda . I takie tam
-
Do pamiętnika .